
汇集公众科学智慧 交流科学思想见解点燃科学智慧火花 构建互动交流平台
汇集公众科学智慧 交流科学思想见解点燃科学智慧火花 构建互动交流平台
一、什么是湖泊热力分层?
“湖泊热力分层”是指湖水温度随深度而变化,呈层状分布的现象。在温暖季节,上层水温较高,下层水温较低,上下层之间水温急剧下降,形成一个突变层,也称为温跃层。在底层湖水水温低于4 °C时,湖泊上层的热量主要由湖水的对流即动力混合来完成。例如,温度为3 °C的水体,表层增温到4 °C时,水的密度最大,因此下沉到深处,同时,密度较小的水上升而产生水质点的对流。这种对流可使水温均达到4 °C为止。
二、根据热力分层特征划分的湖泊类型
多次混合型湖泊(Polymictic):多次混合型湖泊是永久性(连续多次)或经常性(不连续多次)混合的,通常由于其深度较浅,如果经历冬季冰盖,则可被归为冷多次混合型,如果不结冰,则可被归为暖多次混合型。
二次混合型湖泊(Dimictic):二次混合型湖泊每年经历两次混合事件,一次通常发生在夏季分层期之后,另一次发生在冬季逆分层期之后。
单次混合型湖泊(Monomictic):单次混合型湖泊每年经历一次垂直混合事件,通常在冬季。如果它们经历了冬季的冰盖,它们可以被分为冷单次混合型湖泊,如果它们不结冰,它们可以被分为暖单次混合型湖泊。
寡循环型湖泊(Oligomictic):寡循环湖在大多数年份持续分层,但在其他年份完全混合。
局部循环型湖泊(Meromictic):局部循环型湖泊是持续分层的,通常是由于其巨大的深度或化学梯度的存在。
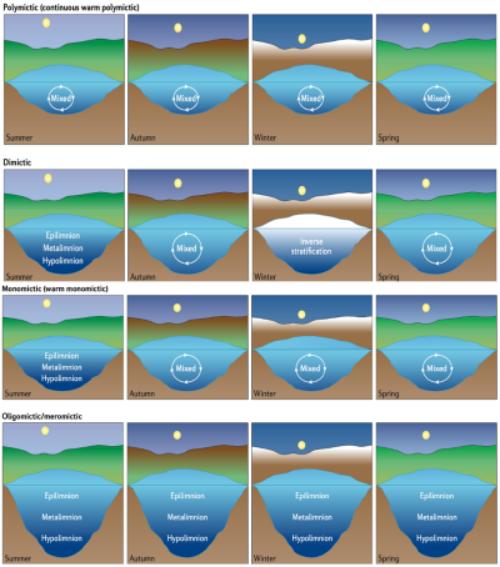
图1 根据热力分层特征划分的湖泊类型
图源:Woolway R I et al., 2020.
三、湖泊热力分层与翻转混合的成因及季节性变化
气温的变化会引起水温改变,而水温的变化会导致湖水密度的不同,因此,大多数温带湖泊在夏季会发生分层现象,而在秋季会发生翻转混合。在寒冷气候下的湖泊中也会发生冬季分层与春天翻转混合的现象。夏季湖泊表层水被太阳辐射直接加热,也被暖空气间接加热,表层水的密度比底层水低,因此表层较暖的水多停留在水面上,直到由于风浪扰动、降水或其他作用力使其向下混合。在分层状态下,表层水由于混合良好而处于好氧状态,底层水因得不到上层水中溶解氧的补充而导致溶解氧较低而处于厌氧状态。表层水的深度与湖泊大小有关,在较小的湖泊中可能仅l m左右,而在大湖中可深至20 m或更深。表层水的深度也与季节性降水有关。在春季表水层正在形成,强降水会使表层较暖的水混合至较深处。表层水一旦形成,湖泊分层会相当稳定,只有较强的风、降水扰动等才可能打破分层状态。
夏季表层水持续变暖而使得热力分层的稳定性不断增加,此时底层水的温度会保持相当稳定。当秋季气温下降,表层水开始冷却,当表层水的密度比底层水高时,表层水便开始下沉,于是发生翻转混合现象,热力分层状态被打破。底层水从而翻转升至湖的表层,在表层被冷却后而下沉。此时,湖泊中发生完全混合。如果气候寒冷,当温度达4 °C时,翻转混合停止,因为4 °C是水的密度最高时的温度。表层水进一步被冷却时,由于表层水的密度会小于深层处4 °C水的密度,于是产生冬季分层现象。当春季来临,湖水开始变暖,湖泊又开始翻转而完全混合。因此,在温带湖泊中每年至少有一次分层与翻转混合现象发生。
四、湖泊热力分层的生态环境影响
气候变化会加剧热分层,减小混合深度,促进有害藻华发生,随之而来的光吸收增加,导致水温升高,形成正反馈,进一步促进了水体热分层。热力分层对水体理化性质、生物地球化学过程和水生态也会产生一系列影响,影响氧浓度,如使得分层的深湖氧浓度降低,减少深水中氧的供给,导致更多甲烷排放和沉积物-水界面的养分释放;然而在冰覆盖湖泊中氧浓度增加。深水湖库往往存在垂向热力分层,决定了溶氧和营养盐等化学因子以及浮游植物和浮游动物等生物因子垂直分层和混合交换,进而深刻影响湖库生态系统结构和功能。
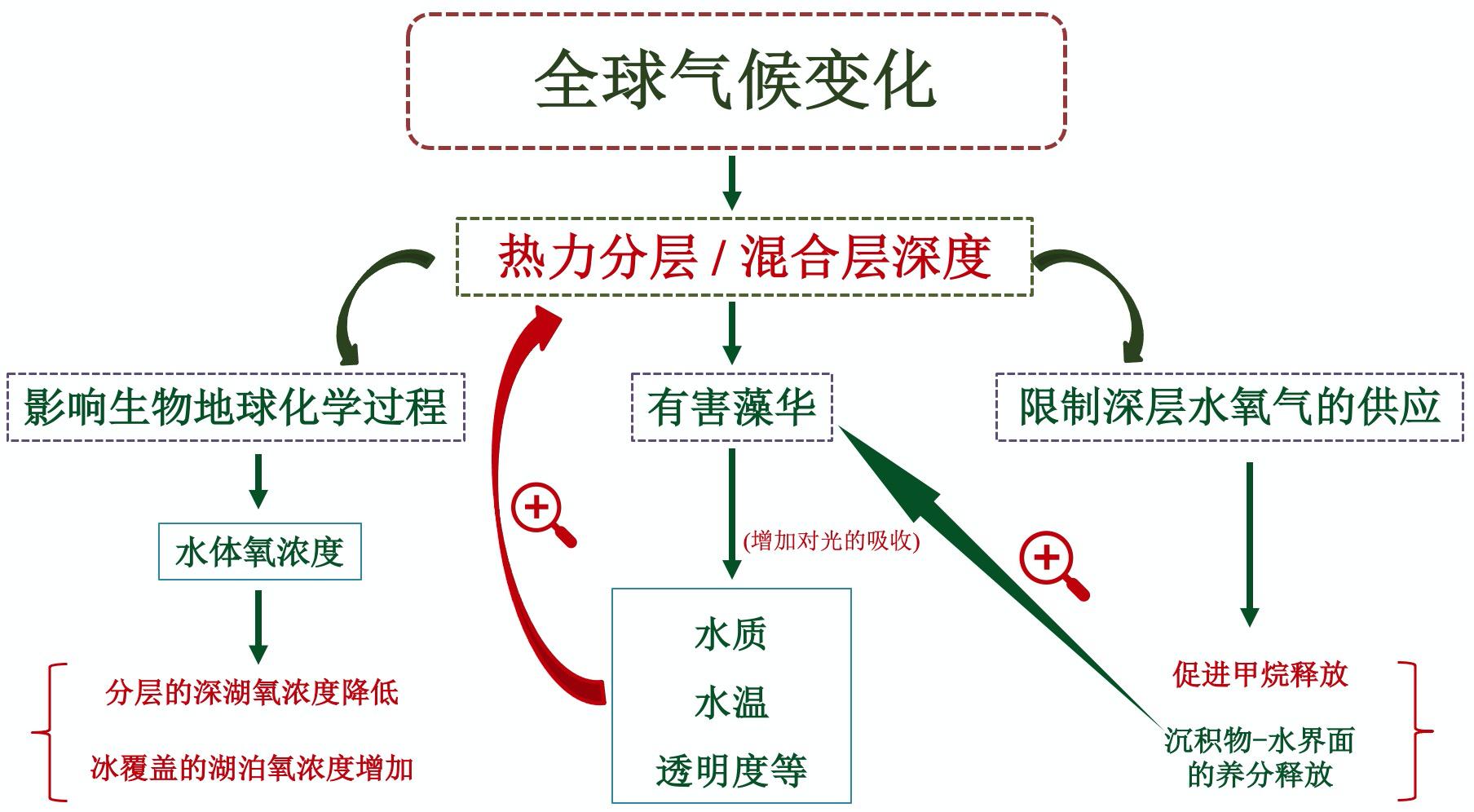
图2 气候变化下的湖泊热力分层及其生态环境效应
空气-水界面的能量通量代表了气候和湖泊之间的相互作用。热分层对于水层之间的传输非常重要,主要由表层较高的水温高于底层形成。光穿透引起表层加热,对浮游植物的生长至关重要。深水混合是强对流冷却的结果,标志着分层季节的结束。风也促进了混合层的混合和加深。冰覆盖影响地表热通量,减少风对湖泊内部的影响。氧浓度与水柱中的许多化学和生物过程有关。营养物质和其他类型的溶质可以从沉积物中释放出来,同样在厌氧环境中沉积物会释放出更多的甲烷等温室气体。如果由于溶质含量的原因,一个湖泊的深水层比上覆水重,则会形成半混合。浮游植物生物量是通过消耗湖泊光照区的营养物和光等资源来生长的,有些蓝藻具有可变浮力,可以从温跃层下吸收营养物质,或者利用浮力在湖中的温跃层中形成深层叶绿素最大值。
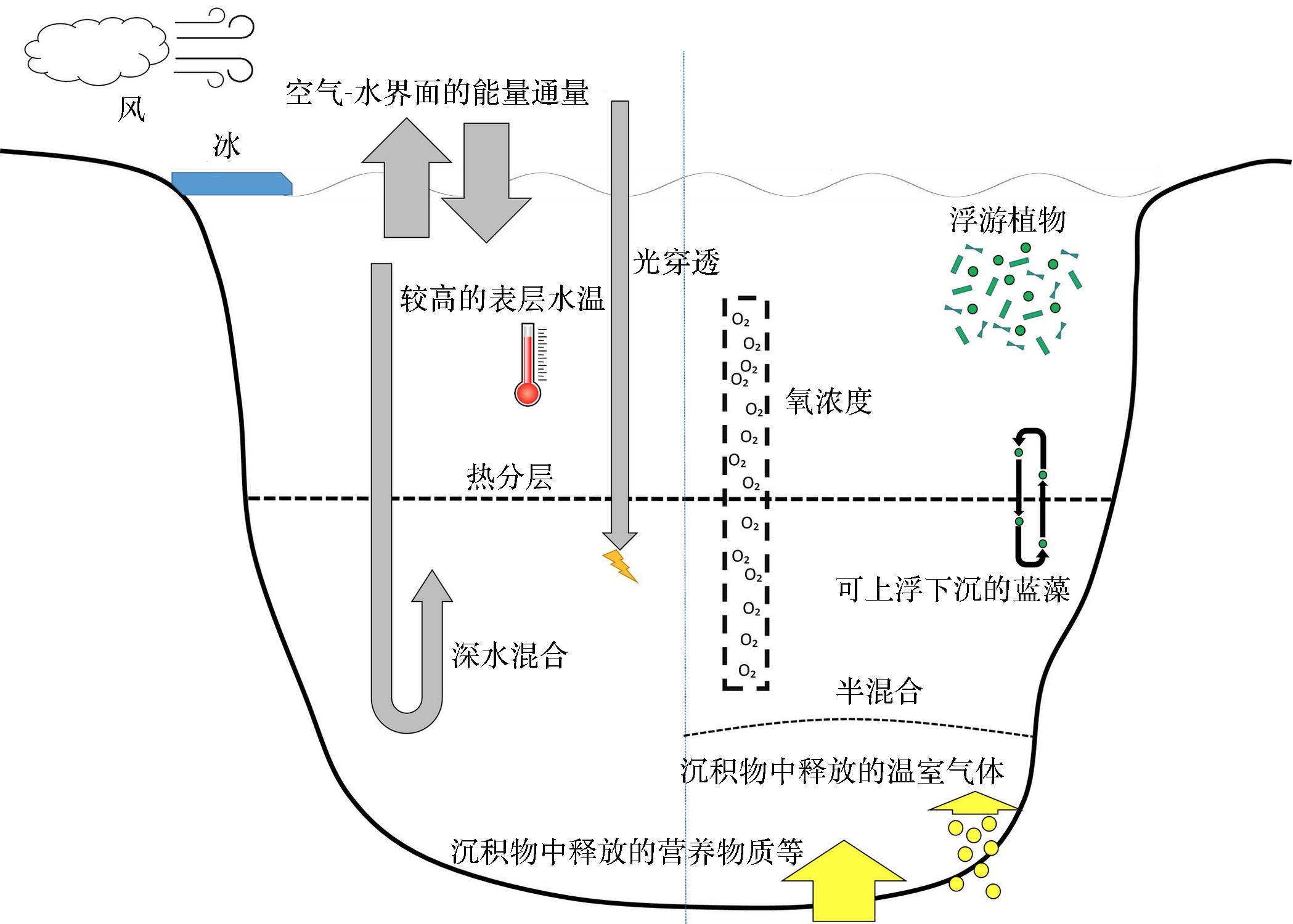
图3 分层的深湖中物理和生物地球化学过程
温度和分层的变化对湖泊生态系统产生了深远的影响。表层水温度升高,更稳定和更长的分层可能有利于形成蓝藻水华并影响湖泊生产力。此外,当湖泊分层时,深层水与大气中的氧气分离,并且分层持续的时间越长,由于湖内呼吸作用而使氧气消耗的越多,导致了缺氧条件和深水死区的形成。这不仅限制了大多数鱼类的栖息地,而且改变了水化学平衡,促进了甲烷的产生。
温度和分层的变化进一步影响生物和地球化学过程。氧浓度可能在深的分层湖泊中降低和在覆冰湖泊中增加。气候变化引起的降水和气温升高的格局变化改变了流域的水文状况,是流域发生各种过程的驱动力。夏季分层的延长会减少向深水的溶解氧补充,加剧缺氧状况。食物网结构的重组和优势种的转移,包括外来入侵物种的增殖,可能是热变化的影响。
温度升高和透明度下降导致的热稳定性增加可能是由于表层和温跃层的水温比下层的水温增加得快。因此,分层时间将延长,混合将减少,这将对湖泊生态系统产生重大不利影响。夏季的分层起着屏障的作用,抑制了水柱的混合。表层水中的温水无法穿过深水层中冰冷、稠密的水。由于水柱的不完全混合和深水层处光合作用的光照不足,水柱可能变得缺氧。此外,浊度通常在温跃层区域达到峰值,这与最大密度梯度的位置密切相关,因此低湍流稳定了垂直水柱中的沉积物,在世界各地的许多水库和湖泊中观察到由于热分层而导致的夏季水质退化。
除了对水质的影响外,分层持续时间的增加也增加了分层对浮游植物功能性组成和群落的影响。
在分层湖泊条件下,浮游蓝藻微囊藻在下沉硅藻和绿藻中占主导地位。这发生在弱混合期间,因为浮游蓝藻物种能够向下迁移以增强其在夜间获取营养的能力,并在白天向上迁移以获取光线,同时形成密集的表面水华并遮蔽其他藻类物种。具有蓝藻水华历史的温暖分层湖泊最容易在未来发生更频繁的水华。蓝藻表面密集的水华可能通过其光合色素和光保护色素对光的强烈吸收而局部提高水温。
温度升高不仅加强了垂直分层,而且降低了水的粘度。根据斯托克斯定律,蓝藻等小生物的垂直速度与粘度成反比。因此,温度升高会降低水体对浮游植物垂直迁移的抵抗力。这将有助于浮力蓝藻形成表层水华,有利于快速垂直调整迁移蓝藻,以优化其养分和光的获取。相反,当粘度降低时,沉降硅藻会遭受更高的沉降损失。这些相反的反应进一步增强了浮蓝藻相对于层状水体中下沉硅藻的竞争优势。当在分层过程中,例如由于浮力调节,浮游植物可利用累积的养分时,随之而来的水华会降低透明度,导致上层水层更强的加热,从而稳定了分层状态。
了解反馈机制的重要性以及湖泊混合状态变化时生物地球化学的作用,可以更好地了解气候变化如何影响湖泊生态系统。强烈的分层和缺氧也会导致湖泊更多的甲烷排放。
大气中二氧化碳含量的增加可能导致湖泊表层水酸化,作为温室气体之一同样会加剧全球变暖。然而,在蓝藻水华密集的系统中,这种影响可能会被水华活动的增加所抵消,而水华活动的增加会增加二氧化碳的消耗,从而提高pH值。热力分层也影响了浮游植物在表层和透光区停留的能力。温跃层的形成减少了浮游植物混合的深度,有效地增加了它们留在光带的机会。
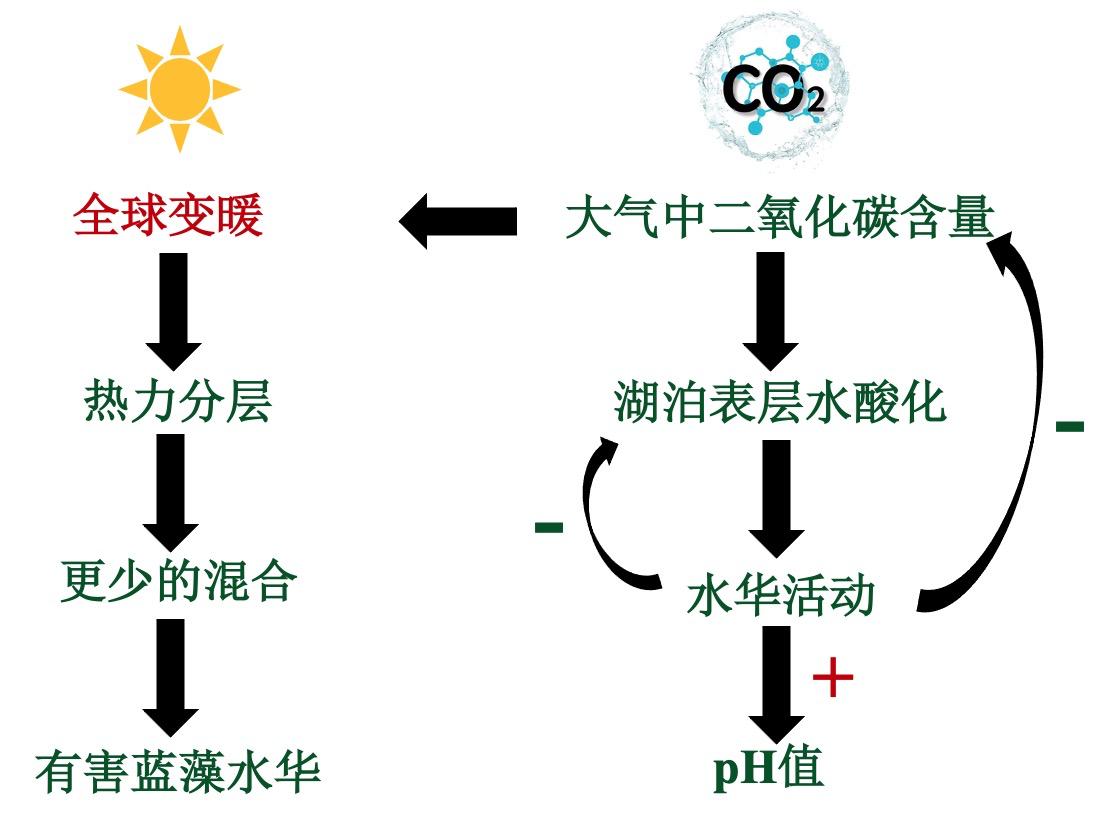
图4 大气中二氧化碳对湖泊热力分层和生态环境的影响
参考文献
《环境科学大辞典》编委会. 环境科学大辞典(修订版)[M]. 中国环境科学出版社, 2008.
Borrel G, Jézéquel D, Biderre-Petit C, et al. Production and consumption of methane in freshwater lake ecosystems[J]. Research in microbiology, 2011, 162(9): 832-847.
Del Giudice D, Zhou Y, Sinha E, et al. Long-term phosphorus loading and springtime temperatures explain interannual variability of hypoxia in a large temperate lake[J]. Environmental science & technology, 2018, 52(4): 2046-2054.
ElciS. Effects of thermal stratification and mixing on reservoir water quality[J]. Limnology, 2008, 9(2): 135-142.
Fang X, Stefan H G. Simulations of climate effects on water temperature, dissolved oxygen, and ice and snow covers in lakes of the contiguous US under past and future climate scenarios[J]. Limnology and Oceanography, 2009, 54(6part2): 2359-2370.
Grasset C, Mendon?a R, Villamor Saucedo G, et al. Large but variable methane production in anoxic freshwater sediment upon addition of allochthonous and autochthonous organic matter[J]. Limnology and Oceanography, 2018, 63(4): 1488-1501.
Huisman J, Sharples J, Stroom J M, et al. Changes in turbulent mixing shift competition for light between phytoplankton species[J]. Ecology, 2004, 85(11): 2960-2970.
Joehnk K D, Huisman J E F, Sharples J, et al. Summer heatwaves promote blooms of harmful cyanobacteria[J]. Global change biology, 2008, 14(3): 495-512.
Matzinger A, Schmid M, Veljanoska-Sarafiloska E, et al. Eutrophication of ancient Lake Ohrid: Global warming amplifies detrimental effects of increased nutrient inputs[J]. Limnology and Oceanography, 2007, 52(1): 338-353.
Merino-Ibarra M, Monroy-Ríos E, Vilaclara G, et al. Physical and chemical limnology of a wind-swept tropical highland reservoir[J]. Aquatic Ecology, 2008, 42(3): 335-345.
Mesman J P, Stelzer J A A, Dakos V, et al. The role of internal feedbacks in shifting deep lake mixing regimes under a warming climate[J]. Freshwater Biology, 2021, 66(6): 1021-1035.
Mullin C A, Kirchhoff C J, Wang G, et al. Future projections of water temperature and thermal stratification in Connecticut reservoirs and possible implications for cyanobacteria[J]. Water Resources Research, 2020, 56(11): e2020WR027185.
O'Reilly C M, Alin S R, Plisnier P D, et al. Climate change decreases aquatic ecosystem productivity of Lake Tanganyika, Africa[J]. Nature, 2003, 424(6950): 766-768.
Regier H A, Holmes J A, Pauly D. Influence of temperature changes on aquatic ecosystems: an interpretation of empirical data[J]. Transactions of the American Fisheries Society, 1990, 119(2): 374-389.
Reynolds C S. The ecology of phytoplankton[M]. Cambridge University Press, 2006.
Schmid M, Hunziker S, Wüest A. Lake surface temperatures in a changing climate: a global sensitivity analysis[J]. Climatic change, 2014, 124(1): 301-315.
Vachon D, Langenegger T, Donis D, et al. Influence of water column stratification and mixing patterns on the fate of methane produced in deep sediments of a small eutrophic lake[J]. Limnology and Oceanography, 2019, 64(5): 2114-2128.
Verburg P, Hecky R E, Kling H. Ecological consequences of a century of warming in Lake Tanganyika[J]. Science, 2003, 301(5632): 505-507.
Wilhelm S, Adrian R. Impact of summer warming on the thermal characteristics of a polymictic lake and consequences for oxygen, nutrients and phytoplankton[J]. Freshwater Biology, 2008, 53(2): 226-237.
Woolway R I, Kraemer B M, Lenters J D, et al. Global lake responses to climate change[J]. Nature Reviews Earth & Environment, 2020, 1(8): 388-403.
Woolway R I, Merchant C J, Van Den Hoek J, et al. Northern hemisphere atmospheric stilling accelerates lake thermal responses to a warming world[J]. Geophysical Research Letters, 2019, 46(21): 11983-11992.
Zhang Y, Wu Z, Liu M, et al. Thermal structure and response to long‐term climatic changes in Lake Qiandaohu, a deep subtropical reservoir in China[J]. Limnology and Oceanography, 2014, 59(4): 1193-1202.
