
汇集公众科学智慧 交流科学思想见解点燃科学智慧火花 构建互动交流平台
汇集公众科学智慧 交流科学思想见解点燃科学智慧火花 构建互动交流平台
朱鹮隶属鹈形目鹮科朱鹮属,是一种中等体型的涉禽,雌雄外型相近,体长大约55-84 cm,主要栖息于温带丘陵和山地森林中,经常在水稻田、湖泊、池塘和草地等环境中活动觅食,以小鱼、泥鳅、蛙、蟹、虾、蚯蚓和甲虫等小型脊椎动物和无脊椎动物为食。朱鹮为一夫一妻制鸟类,除非一方发生意外,否则极少改变配偶。每年的活动周期大致可分为三段,分别为繁殖期(2月-6月)、游荡期(7月-10月)和越冬期(11月-2月)。其每年繁殖一次,若第一次繁殖失败较早,也可能进行第二轮繁殖。亲本通常轮流坐巢,共同完成孵卵及育雏工作。至越冬期,朱鹮会逐渐聚集成群共同觅食和夜宿。

图1. 朱鹮在池塘觅食 (席学博 摄于河南董寨)
历史上,朱鹮曾广泛分布于东亚地区,从中国台湾到俄罗斯西伯利亚,从日本到中国甘肃、青海均有其活动记录。但由于栖息地环境恶化、猎捕等因素影响,其种群数量急剧下降,在20世纪中后期曾一度被认为已经灭绝,直到80年代,中国科学院动物研究所刘荫增先生等研究人员才在陕西省洋县发现了唯一的野生种群 (7只个体,有3只为其中一个繁殖对的后代)。

图2. 朱鹮历史分布图 (Li et al., 2009, Biodiversity and Conservation)
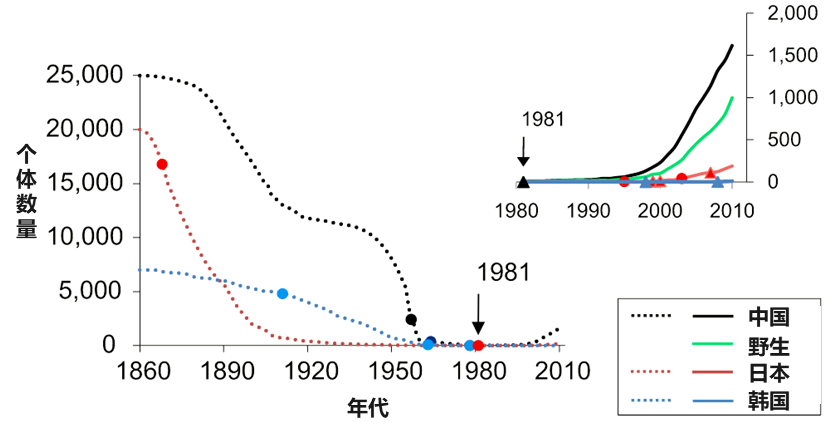
图3. 朱鹮种群数量变化情况 (Li et al., 2014, Genome Biology)
自此,我国政府将朱鹮列为“中国濒危珍稀动物”、“国家一级重点保护野生动物”,同时它也被国际列入“濒危野生动植物种国际贸易公约附录物种名录”和世界自然保护联盟IUCN“濒危物种红色名录”中。目前,通过实施就地保护、人工繁育和再引入等各种强有力的保护措施,全球朱鹮种群数量已复壮到1万余只。在中国陕西、河南及日本和韩国等地都已经建立了人工与野生种群,朱鹮的栖息地面积也由当初发现时的不足5平方公里,增长至1.6万平方公里。朱鹮保护工作因此成为了拯救濒危物种的成功典范,与大熊猫、亚洲象和海南长臂猿一同作为保护成果写入中国生物多样性保护白皮书。
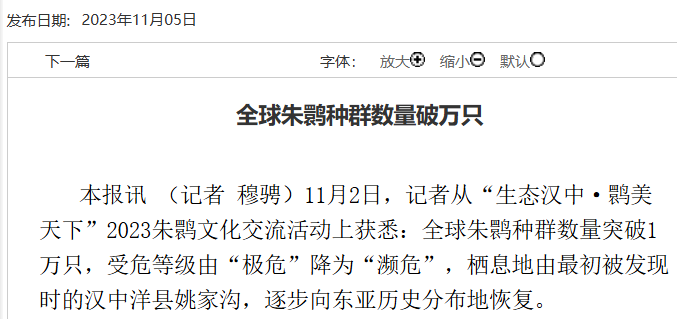
图4. 2023年全球朱鹮种群数量破万只 (来源:陕西日报,https://esb.sxdaily.com.cn/pc/content/202311/05/content_846774.html)
目前,关于朱鹮的研究相对集中于基础的行为描述、繁殖扩散和栖息地偏好等工作中,也有部分关于遗传和基因组方面的研究进展。但相比于其他明星物种,如大熊猫,朱鹮还是比较缺乏长期的基础机制研究。尽管近些年朱鹮一直处于种群恢复增长的趋势中,但是我们并不清楚这种增长背后的生物学机制。同时,野生朱鹮种群仍相对集中于陕西省,野外分布情况尚未完全恢复到到历史水平,一旦出现疫病爆发和其它极端自然灾害等意外,朱鹮种群极有可能遭受灭顶之灾。因此,这个物种仍处于“濒危”状态,亟需我们的关注与重视。
传统的保护生物学理论和观点认为:一个物种的有效种群数量 (即能正常繁殖的个体数)如果特别小,那么它将面临近交繁殖以及遗传瓶颈效应等问题。所谓遗传瓶颈效应,就是指种群像经过瓶子的颈部一样,数量急剧下降,而其原本拥有的遗传变异因此大量丢失。这会导致种群的遗传多样性下降、有害突变累积增加,其数量理论上很难恢复到正常水平。然而,朱鹮种群的变化过程却并非如此,尽管最后的野外有效种群数量仅为两个繁殖对(四个成年个体),该极端小种群如今已发展复壮到了可观水平,这与传统保护生物学理论所预测的情况并不符。朱鹮究竟如何创造了这样的奇迹,其背后的机制仍然需要更多的研究。
为了解答这个问题,我们正在河南董寨国家级自然保护区开展系统的研究工作。河南董寨国家级自然保护区位于河南省信阳市罗山县境内,地处大别山西段,气候宜人、四季分明、植被类型丰富,是众多鸟类的栖息和繁衍场所,也是许多候鸟迁徙过程中的重要停歇地和栖息繁衍地。2004年,董寨保护区被确定为中国陕西省外第一个朱鹮再引入野化放飞点。2007年起,保护区以来自北京动物园的4只和日本返还的13只朱鹮为奠基者,开始人工繁育和野化放飞工作。截至2023年,由保护区人工繁育长大的个体累计已有约300只,同时已有7批共133只朱鹮经过野化训练后被放归到野外自由繁殖生活。据估计,目前河南董寨保护区及周边地区的野外朱鹮种群数量已有400余只,种群结构也呈现为稳定增长的“金字塔”型 (幼年个体多、老年个体少),其发展趋势整体稳中向好。

图5. 董寨朱鹮繁育中心
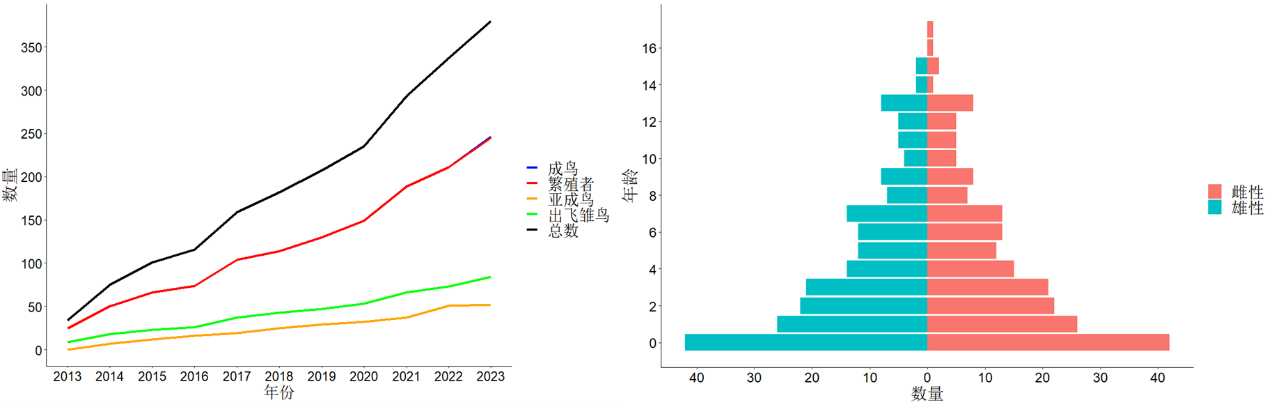
图6. 董寨野外种群数量(左)与种群结构(右)估计情况
为了探究朱鹮种群如何挣脱小种群的限制,我们计划从以下几个方面入手解决这个问题。
首先,我们需要追踪董寨种群建立以来每个个体的命运,并且梳理清楚整个种群的家系,也就是个体间的亲缘关系情况。除了参考历史繁殖记录,我们也通过采集血液和羽毛等样品进行测序鉴定,从而获得更准确的基因与家系信息。有了这张关系网,我们就能知道每个个体活了多久、是否健康、与谁配对、有多少后代,同时也就能计算出重要繁殖参数 (比如不同繁殖配对的窝卵数、卵受精率、胚胎死亡率和雏鸟出飞率等) 的遗传力,知道它们在多大程度上受遗传因素的影响。了解重要性状的遗传进化将有助于我们掌握这个朱鹮种群的发展历史;定量近交繁殖程度及其对种群的影响;评估种群遗传多样性;以及为优质性状、优质个体的选育提供参考。
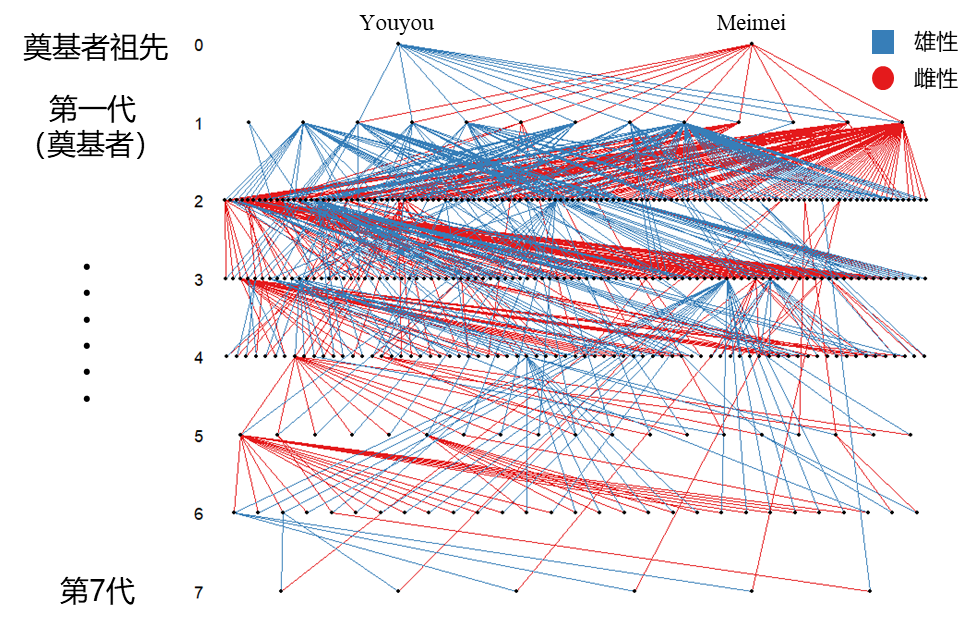
图7. 董寨人工繁育种群家系图 (基于保护区15年间的繁育数据)

图8. 工作人员对朱鹮进行采样与身体数据测量 (何远思 摄于河南董寨)
除了遗传,我们也关注朱鹮配偶行为相容性对其繁殖结果的影响。
配偶间行为相容性在进化和保护等领域都具有重要的意义与潜在应用前景。研究表明其与物种对环境压力的抵抗能力和适应性有关,也对笼养种群的繁殖能力及成功率有一定影响。行为间的不相容可能导致雌雄个体间繁殖合作不佳,以及出现频繁的配对内攻击行为,使配对双方因为难以和谐相处而无法完成交配,甚至造成个体死亡,给物种保护工作带来损失。据报道,日本最后的朱鹮便是因为配对双方行为不相容,在繁殖季出现激烈冲突,以至配对失败,最终朱鹮在日本本土灭绝。因此,完善对朱鹮配偶行为相容性的研究将有利于我们进一步理解物种濒危的原因,并制定更具有针对性的保护措施。
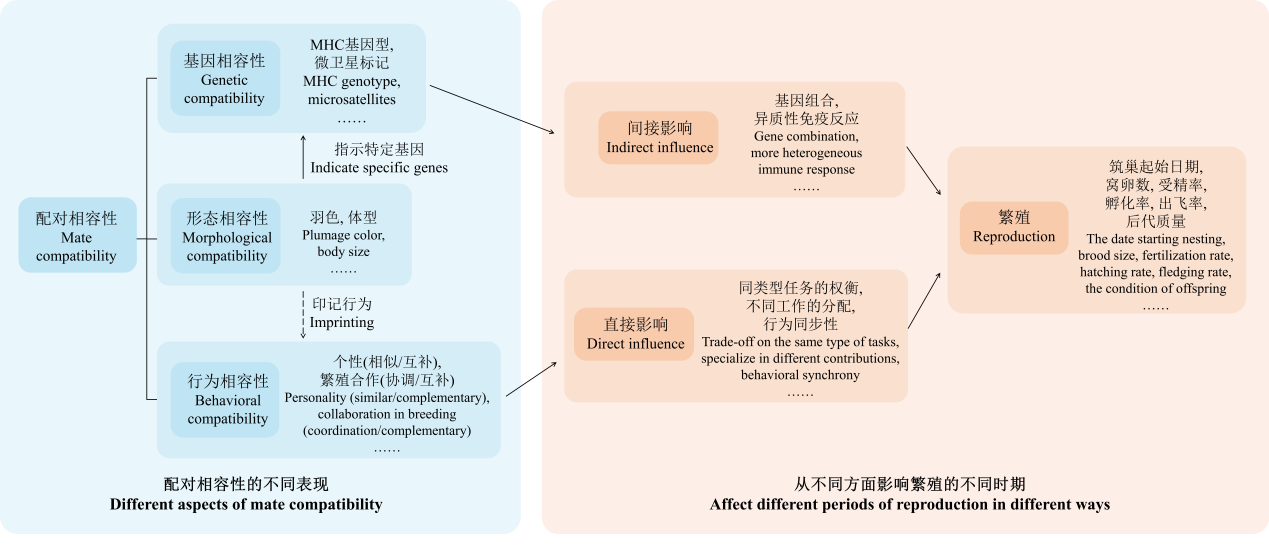
图9. 配对相容性对繁殖的影响 (何远思 等, 2023, 生物多样性)
为了收集相关数据,除了借助摄像机记录野外朱鹮的繁殖情况,我们还在董寨保护区的荒田保护站内建立了10个半自然繁殖笼。这些繁殖笼都位于远离道路的地方,并且除了定时喂食与清扫水池外,我们尽可能不打扰已配对的朱鹮,以保证它们在最接近自然状态的情况下生活,所有卵都由朱鹮自孵自育,除非特殊情况 (比如父母不会喂食或被捕食等),否则都尽量不干预它们。同时,我们通过监控观察记录其行为,对雌雄个体在孵化和育雏工作方面的参与情况以及个体间的亲密程度进行定量,通过比较不同配对的行为以及繁殖成功率的差异探究配偶间行为相容性对朱鹮繁殖的影响,以期更深入理解影响朱鹮繁殖成功率的因素,并帮助我们判断哪些配对个体相处得更好、什么样的行为表现代表雌雄双方关系不佳,从而为人工配对选育提供参考,提高繁育工作效率。

图10. 安装的摄像机(左)与摄像机监测画面(右)

图11. 半自然繁殖笼(左)与繁殖笼监控画面(右)
在此基础上,我们得以着手探索我们最关心的问题——朱鹮种群濒危与复壮的机制。
基于此前所得的家系、遗传数据、通过计算繁殖对之间的近交程度与其繁殖成功率相关指标间的关系,我们能科学定量朱鹮种群的近交程度以及其对种群增长的影响情况。进一步地,通过联系朱鹮的配偶选择、亲本繁殖行为和越冬集群情况等数据,我们能探究梳理该物种是否存在潜在的避免近交繁殖与衰退的关键遗传与行为特征,从而在性选择和自然选择理论的框架下;了解朱鹮种群在应对小种群限制和环境压力时发展出的适应性策略,揭示朱鹮种群的变化历史以及这个物种能成功恢复的原因;最终明晰其濒危与复壮的生物学机制,并制定更具针对性的保护策略。

图12. 秋冬季朱鹮集群夜宿 (席学博 摄于河南董寨)
对朱鹮进行研究的意义不仅在于保护这一珍稀濒危物种,更重要的是,一方面朱鹮可以作为典型的保护案例,为我们理解其他珍稀濒危物种的濒危原因、制定保护计划提供借鉴与参考;另一方面,围绕朱鹮进行的保护工作也有利于对其分布区内整体生态系统功能及生物多样性的保护,其作为“明星”物种所具有的示范效应,能有效吸引公众力量参与保护,并进一步起到辐射带动作用。
