
汇集公众科学智慧 交流科学思想见解点燃科学智慧火花 构建互动交流平台
汇集公众科学智慧 交流科学思想见解点燃科学智慧火花 构建互动交流平台
我在动物研究所分会以我国农药残留标准体系建设进展与挑战科学沙龙之后写了一篇“农药残留何时休”的文章,很多人不知道什么是植物免疫,甚至认为植物还有免疫吗?下面就讨论一下植物免疫,特别是活性氧和一氧化氮自由基在植物还有免疫中的作用。
一、何为植物免疫
动物或人免疫是指体内的抗体能够通过淋巴液和血液循环系统遍布全身,而植物没有血液循环系统,那么它的抗病能力是怎么遍布所有组织的呢?和动物类似,当病原微生物侵染植物时,植物能通过细胞膜表面受体识别病原的侵入,例如病原的鞭毛蛋白的某些氨基酸通过识别这种病原应对大部分的病原微生物信号时,会把病原信号传递给细胞核,之后植物做出一系列反应,例如将气孔关闭,将细胞壁加厚,合成溶菌物质等增强植物对病虫害的抵抗能力。因此,植物免疫剂对减少甚至免除农药具有重要意义 [1]。
1、模式触发免疫(PTI)和效应子触发免疫(ETI)
植物免疫系统由两个主要层次构成:PTI和ETI。PTI通过识别多糖、鞭毛蛋白等病原保守分子模式触发基础防御反应,通过细胞膜表面的模式识别受体(PRRs)感知病原相关分子模式,激活钙离子内流。ETI则依赖植物R蛋白特异性识别病原效应蛋白引发更强烈的抗性反应。植物免疫剂可以通过激活植物调节自身代谢来增强免疫系统,赋予植物抗病能力 [2]。
2、植物免疫信号由防御激素水杨酸(SA)增强
免疫信号由防御激素SA增强,通过重编程转录组来促进防御。SA类型免疫受体激活机制和SA感知信号传导与不同受体组织成网络,整合复杂信号产生适当防御输出[3]。PRRs的激活会提高防御基因表达和植物激素SA的积累,介导下游免疫反应,为植物提供对病原体的强效抗性[4]。
3、钙离子Ca²⁺在真核生物免疫中充当关键的第二信使
Ca²⁺在植物免疫中发挥关键作用,是所有真核细胞中重要的细胞内信使。Ca²⁺通过离子通道、交换器和泵的协同作用,在空间上和时间上特定的胞质Ca²⁺信号得以形成,以响应各种刺激;这些信号随后通过Ca²⁺结合感应蛋白的作用被解码。Ca²⁺信号对于PTI和ETI免疫都至关重要 [5]。Ca²⁺可渗透通道被激活。近期研究发现,Ca²⁺信号在由质膜和细胞内受体分别启动的免疫反应两个层面中都起着核心作用[6]。
二、活性氧(ROS)和一氧化氮(NO)在植物免疫系统发挥重要作用
快速产生的ROS和NO被认为在植物免疫调控中发挥作用。
1、活性氧(ROS)
ROS在植物信号传导中起核心作用。植物免疫中ROS生成与信号传导通过复杂机制。微生物诱导的胞间隙ROS生成调控与脱落酸(ABA)诱导的胞间隙ROS发挥免疫作用。ROS作为信号分子调控细胞活动,并以气孔运动结合发挥作用[7]。ROS信号在植物免疫反应中主要由 NADPH 氧化酶介导。植物细胞表面定位的模式识别受体(PRRs)能够识别微生物并激活模式PTI[8]。磷脂酸(PA)和ROS是介导多细胞生物和植物中各种信号过程的重要细胞信使。PA 能够结合并稳定 NADPH 氧化酶,调节植物PTI和ETI中的 ROS 生成及其增强。细胞PA爆发在协调植物免疫中调控ROS的生成[9]。
植物细胞死亡在植物免疫中由钙、铁和ROS信号介导。超敏反应(HR)参与铁死亡。铁死亡是一种受调控的、非凋亡的依赖铁和ROS的植物细胞死亡形式。病原体感染会触发植物细胞内铁的积累和ROS的骤增,钙、铁和ROS信号在植物免疫过程中铁死亡中发挥关键作用 [10]。PTI和ETI两者都利用Ca²⁺内流和ROS进行信号传导触发细胞死亡机制,限制病原体扩散。Ca²⁺和ROS的这种复杂利用强调了NLR在植物先天免疫中弥补适应性免疫系统缺失方面的关键作用[11]。
2一氧化氮(NO)
NO在植物防御真菌病原体的调控中具有重要作用。NO 会触发防御相关基因表达的重编程产生以及超敏反应。NO 在植物-真菌互利共生关系建立的早期阶段也具有调控作用。在识别根部真菌共生体后,植物会产生 NO [12]。NO调节着一系列免疫反应,激活一组与防御相关的基因及水杨酸依赖性植物免疫的正向调控因子,能够调控关键的免疫相关氧化还原信号 [13]。
3一氧化氮(NO)和活性氧(ROS)的协同作用
NO和ROS在防御反应中协同作用。NO和ROS的快速生成已被认为参与植物的多种生理过程,如程序性细胞死亡、发育、细胞伸长和激素信号传导。这些信号分子在植物先天免疫中的调控作用发挥重要作用。NADPH氧化酶负责病原体应答的ROS爆发。许多研究表明植物中存在类似NO合酶的活性,能够引起NO的突发生成。由丝裂原活化蛋白激酶级联反应、钙依赖性蛋白激酶或核黄素及其衍生物调控NO和ROS爆发的机制在植物抵御病原体反应中的作用[14-15]。
三、如何提高植物免疫
通过施加SA、NO和转基因可以提高植物免疫。
1、通过施用水杨酸(SA)提高植物免疫
激素SA)能够将病原信号从染病位点传递给植物远端的健康组织,当病原侵染其他健康组织时,植物能够大量迅速的调动植物资源快速合成抗病蛋白从而保护植物免受伤害。叶面施用SA能够调控并诱导内源性NO和ROS,从而提高番茄对尖孢镰刀菌胁迫的先天免疫力和活力。在叶面施用SA后,发现SA处理组的病害发生率也显著降低。同时NO生成显著增加,并且与ROS生成与NO之间的直接相关性。用NO合酶抑制剂或NO清除剂联合处理的番茄植物,所有检测的参数显著降低。SA处理的番茄种子还表现出较高的幼苗活力指数、茎和根长度等。SA与内源NO之间的相互作用具有显著提高番茄植株防御能力和生长的潜力[16]。
2、通过施用NO提高植物免疫
单独活性氧还不足以导致细胞死亡,NO介入和协调才可引起植物感染部位细胞的死亡,从而引起过敏坏死反应,二者发挥免疫作用。条锈菌是世界上许多地区小麦的重要病害。50年代以碧蚂一号抗锈性丧失使小麦减产。我们通过喷洒硝普纳(SNP)提高小麦免疫力,可以有效杀伤小麦条锈病,取得了很好的效果。硝普纳产生一氧化氮渗透小麦叶组织后接种致病小种条中29号后,随外源SNP剂量的增加和诱导时间的延长叶面坏死呈现减小的总趋势。如果加入一氧化氮清除剂CPTIO就没有效果。图1 是外源SNP诱导时间和诱导剂量对抗病性症状产生的影响和统计数据结果[17-20]。

图1 外源一氧化氮SNP诱导时间和诱导剂量对抗病性症状产生的影响(左图)和统计数据结果(右图)。
3、通过转基因增加植物免疫
拟南芥转基因SRG2 和 SRG3 过表达株系表现出细胞死亡增加、ROS积累以及持续的致病相关基因1(PR1) 表达。高水平的 SRG2 和 SRG3 过表达导致微小病变的形成,(图 2a)。细胞死亡相对强度微小病变的形成随着 SRG2 或 SRG3 表达量的增加而增加(图 2b)。SRG2和 SRG3株系相对于 SRG2株系表现出更多的细胞死亡(图 2b)。ROS的形成是一个关键的早期防御反应。与野生型Col-0相比,SRG2和SRG3系统均显示出DAB和NBT染色增强(图2c–f)。此外,增加SRG2或SRG3的表达导致DAB和NBT染色进一步增强(图2d,f)。检测基因PR1的表达表明,与野生型Col-0相比,SRG2和SRG3株系中PR1的表达显著增加(图2g)。结果显示,过表达SRG2或SRG3会激活多种免疫反应,包括细胞死亡发生、ROS产生以及PR1表达[21]。
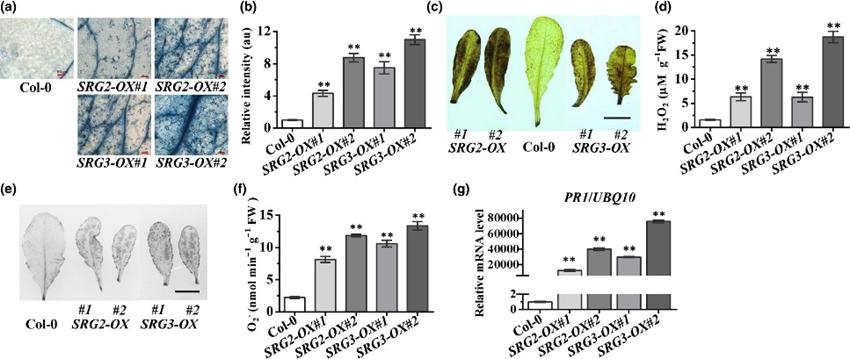
图2 SRG2 和 SRG3 的过表达促进拟南芥关键防御响应的激活。(a) 通过台盼蓝 (TB) 染色评估细胞死亡。 (b)对 TB 染色的相对强度进行分析。 (c) 通过 3,3′-二氨基联苯胺 (DAB) 染色检测植物叶片中 H2O2 的积累。 (d) 对拟南芥幼苗中 H2O2 进行定量 (e) 通过硝基蓝四唑氯化物 (NBT) 染色检测植物叶片中超氧化物的积累。 (f)对超氧化物产生进行定量。(g) SRG2和SRG3株系中PR1的表达显著增加。
参考文献
